André Luis Klein e Claiton Martins-Ferreira
 O objetivo deste artigo é sintetizar conceitos-chave sobre especiação e servir de auxílio para professores e alunos de ciências biológicas. Adotou-se uma abordagem que estimula o questionamento como forma de interligar diferentes temas. Partindo-se dos conceitos de espécie, são tratados: a especiação alopátrica, o isolamento reprodutivo como um subproduto da divergência, o papel da seleção natural, reforço, hibridização, o papel da seleção sexual, especiação simpátrica, especiação parapátrica e mecanismos de isolamento.
O objetivo deste artigo é sintetizar conceitos-chave sobre especiação e servir de auxílio para professores e alunos de ciências biológicas. Adotou-se uma abordagem que estimula o questionamento como forma de interligar diferentes temas. Partindo-se dos conceitos de espécie, são tratados: a especiação alopátrica, o isolamento reprodutivo como um subproduto da divergência, o papel da seleção natural, reforço, hibridização, o papel da seleção sexual, especiação simpátrica, especiação parapátrica e mecanismos de isolamento.
Quantas espécies?
Na Figura 1, em um primeiro momento, quantas espécies de borboletas pode-se dizer que existem? Intuitivamente, as diferenças no padrão de cores das asas parecem justificar a classificação em várias espécies. Porém, são todas subespécies (ou raças geográficas) de Heliconius erato, uma borboleta que se distribui do Uruguai até a América Central (SHEPPARD et al., 1985). Tais espécies com várias formas alternativas são denominadas “politípicas”.
As aranhas da Figura 2 – Paratrechalea ornata e Paratrechalea azul – por outro lado, convivem lado a lado, nos mesmos ambientes, e não apresentam qualquer diferença visível a olho nu que permita identificá-las, podendo ser discriminadas apenas com o auxílio de um estereoscópio por um especialista, com base na estrutura de suas genitálias (COSTA-SCHMIDT; ARAÚJO, 2010). Este é um exemplo oposto do que vimos anteriormente, nas borboletas, pois duas espécies compartilham uma mesma forma, caso em que são chamadas de espécies “crípticas”.

Figura 1.
Borboletas Heliconius erato – um exemplo de “espécie politípica”. Os padrões de coloração das raças geográficas são tão distintos quanto aqueles vistos entre espécies diferentes.
(Extraído de SHEPPARD et al. 1985).
Se é possível haver formas tão diferentes dentro de uma mesma espécie, e espécies diferentes podem ser tão semelhantes, o que significa ser uma espécie? Nestes dois organismos, assim como em muitos outros, a capacidade de reprodução é muito mais importante do que a própria morfologia.
As subespécies de Heliconius erato podem se intercruzar e produzir descendentes férteis, enquanto que as duas espécies de Paratrechalea, não. Sob essa perspectiva pode-se dizer que “espécies são grupos de populações naturais que intercruzam e estão reprodutivamente isoladas de outros grupos desse tipo” (MAYR, 1963). Este é o Conceito Biológico de Espécie (CBE).
Alternativas ao CBE
Pode-se fazer alguma objeção ao conceito biológico de espécie apresentado acima? Sim; as bactérias, por exemplo, não fazem sexo. Além disso, apresentam pouca diversidade morfológica. Como elas se enquadram nestes critérios? Antigamente, para se distinguir bactérias em forma de bastão, por exemplo, elas eram testadas quanto ao seu metabolismo. Um tipo, classificado posteriormente como Escherichia coli, podia se alimentar de lactose, ao passo que outro, classificado posteriormente
como Vibrio cholerae, não. Esta é a lógica do Conceito Ecológico de Espécie – elas são definidas com base nos recursos que são adaptadas a utilizar. Mais precisamente, sob a ótica deste conceito uma espécie é uma linhagem que evolui independentemente e que ocupa uma zona adaptativa minimamente diferente das demais. Atualmente, trabalhos de comparação genômica chegaram à conclusão de que bactérias e arqueobactérias devem ser classificadas com base no seu conteúdo genético. A lógica, contudo, é a mesma da anterior, uma vez que linhagens evolutivamente independentes devem formar agrupamentos genéticos distintos.
E os que trabalham precisamente com o diagnóstico de espécies, isto é, os taxonomistas, o que têm a dizer? Embora hoje a taxonomia tenha incorporado fortemente o pensamento filogenético nas suas bases teóricas, por muito tempo este campo seguiu a lógica do Conceito Fenético de Espécie, ainda utilizado em conjunto com as novas abordagens. De acordo com este conceito as espécies são definidas com base em caracteres observáveis e mensuráveis dos organismos. De fato, na prática do
dia-a-dia reconhecemos as espécies por seus caracteres fenéticos, com maior ou menor sucesso. O conceito fenético se assemelha aos conceitos morfológico e tipológico, quando existe um indivíduo “tipo” que apresenta as características definidoras da espécie.
Como surgem as espécies?
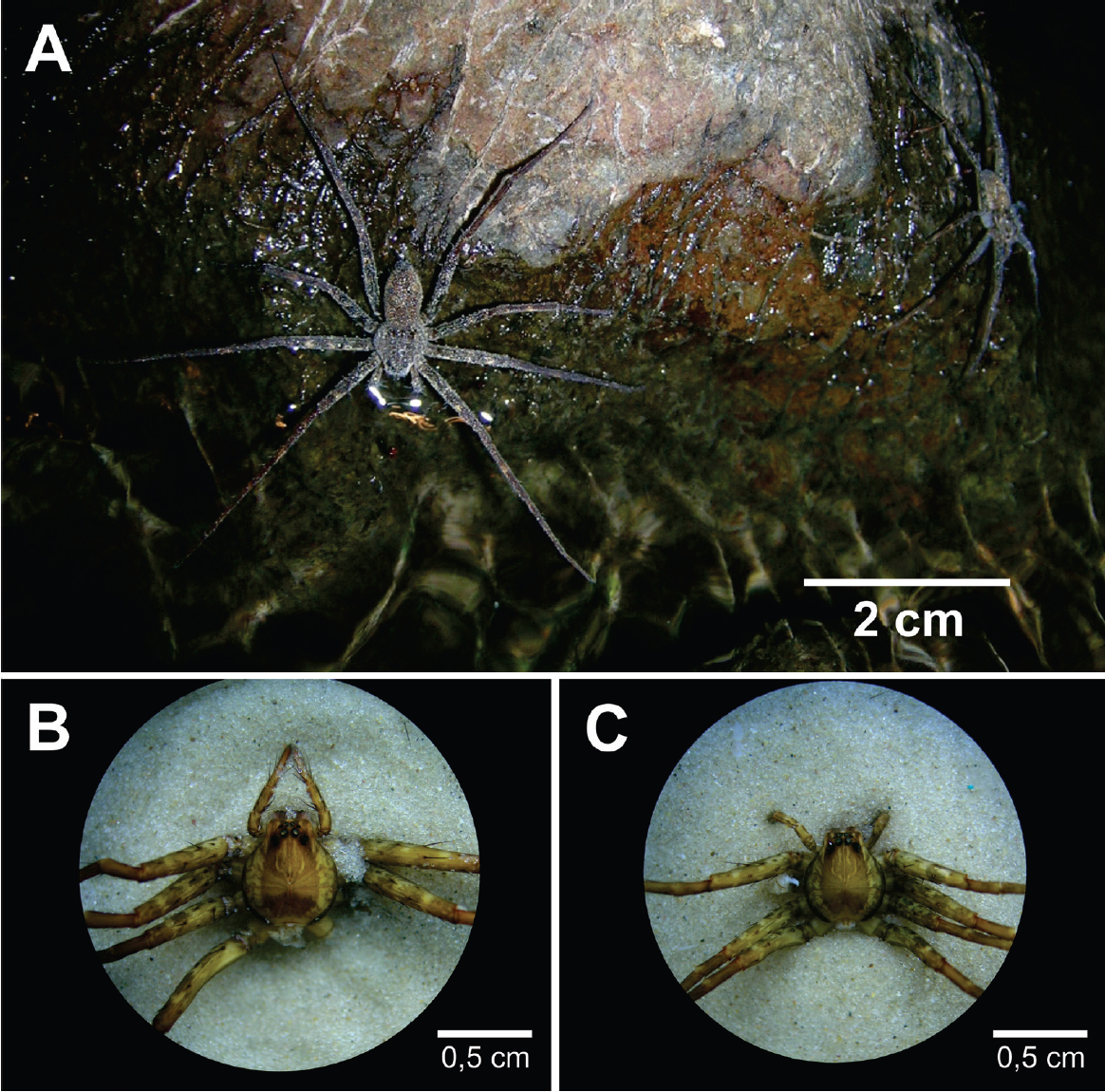
Figura 2.
Aranhas do gênero Paratrechalea – exemplo de “espécies crípticas”. A identificação só pode ser feita por um especialista através da visualização da genitália em estereoscópio. A) P. azul, mais à
frente, e P. ornata, ao fundo. B) Vista dorsal de P. azul; C) Vista dorsal de P. ornata. (Fotos de L.
E. Costa-Schmidt).
Parte da discussão sobre espécies, entretanto, não gira em torno da prática, mas do significado biológico das espécies. Afinal, como elas surgem? Se retomarmos o CBE, vemos que as espécies devem estar “reprodutivamente isoladas” de outros grupos desse tipo. Portanto, a pergunta que devemos fazer é: o que leva as populações a se isolarem reprodutivamente?
Talvez a primeira resposta que nos venha à mente seja “uma barreira física”, como montanhas, rios, vales ou desertos que isole populações da mesma espécie. Podemos pensar na formação do istmo do Panamá, que separou o oceano Pacífico do mar do Caribe há cerca de 3 milhões de anos. Tornou-se impossível populações de espécies aquáticas dos dois lados do istmo se encontrarem. E o que acontece depois? Entra, então, em jogo, o acaso das mutações genéticas. Não que antes isso não ocorresse. Acontece, porém, que agora as mutações de um lado não podem ser disseminadas para as do outro e as diferenças se acumulam. Assim, inicia-se a divergência entre as populações geograficamente isoladas. Se, eventualmente, tais populações voltarem a se encontrar, é possível que os dois conjuntos de genes voltem a formar um só pelo intercruzamento dos indivíduos. Contudo, se as diferenças acumuladas forem tão grandes a ponto de os indivíduos provenientes de um lado não serem mais capazes de cruzar com os do outro, os dois conjuntos genéticos serão mantidos. É importante ressaltar então que apenas com um mecanismo intrínseco (biológico) de isolamento, e não geográfico, é que as populações estão reprodutivamente isoladas e podemos dizer que existem duas espécies. Quando a especiação ocorre em lugares diferentes – nos dois lados do istmo do Panamá, neste caso – diz-se que ela é alopátrica (alo = diferente; patria = lugar) (Figura 3A).
Evidências das etapas da especiação alopátrica podem ser vistas tanto na natureza quanto em laboratório. Populações da borboleta Heliconius erato phyllis no Rio Grande do Sul, por exemplo, apresentam-se em estrutura de “ilhas”, com maior diferenciação genética entre elas do que dentro delas (SILVA E ARAÚJO, 1994). A deriva genética e a baixa capacidade de dispersão desta espécie devem ter sido os responsáveis por essa divergência entre populações geograficamente afastadas, divergência essa que, conforme vimos, pode ser o primeiro passo de um processo de especiação.
Uma evidência de laboratório provém de populações de Drosophila pseudoobscura mantidas isoladas em meios de cultura diferentes – um, à base de amido, e outro, à base de maltose. Após várias gerações, as populações desenvolveram diferenças nas enzimas digestivas e, quando colocadas em contato, os indivíduos preferiam cruzar com aqueles criados no mesmo meio de cultura – ou seja, havia sido iniciado um processo de isolamento reprodutivo desencadeado pelo tempo que as populações permaneceram sem contato em ambientes diferentes (DODD, 1989).
O papel da Seleção Natural
O simples fato de duas populações estarem separadas geograficamente pode causar diferenças genéticas entre elas, por deriva. E a seleção natural, qual seu papel? A resposta está no segundo exemplo a respeito das populações de Drosophila pseudoobscura, onde outro importante fator entrou em jogo: a adaptação. Juntamente com as diferenças adaptativas nas enzimas digestivas, surgiram diferenças importantes, ainda que desconhecidas, no contexto reprodutivo, responsáveis pelas preferências sexuais manifestadas após a divergência. As diferenças que levaram ao isolamento reprodutivo podem ter se originado por pleiotropia (um gene influenciando mais de um fenótipo) ou pelo efeito carona (genes selecionados juntos devido à sua ligação física).
Os famosos tentilhões de Darwin propiciam-nos um exemplo ainda mais claro. Após a dispersão de uma espécie ancestral para as diferentes ilhas de Galápagos, a seleção
natural favoreceu, em cada população, a forma de bico mais adequada para utilizar os alimentos encontrados no seu hábitat. Acontece que bicos não servem só para comer. Pesquisadores constataram que a vocalização de cortejo destas aves era diferente e se relacionava com a morfologia do bico. A atração de parceiros, portanto, foi afetada pela evolução adaptativa para alimentação, levando ao isolamento reprodutivo entre as populações de cada ilha (WEINER, 1995).
Um terceiro exemplo retorna às borboletas que já mencionamos neste artigo: as Heliconius. As diferentes subespécies são, na verdade, mímicos de outra espécie do gênero (H. melpomene). Em cada local, talvez devido a uma fragmentação do ambiente no passado, as populações evoluíram um padrão de coloração próprio que serve como defesa contra predadores. Mas, as cores não têm apenas função protetora nestas borboletas. O reconhecimento de fêmeas pelos machos é feito através das cores das asas, de modo que os machos preferem acasalar com fêmeas do seu próprio padrão. A divergência causada por uma adaptação de defesa, portanto, teve como subproduto um grau inicial de isolamento reprodutivo entre as subespécies (JIGGINS, 2008).
Conceitos mais debatidos
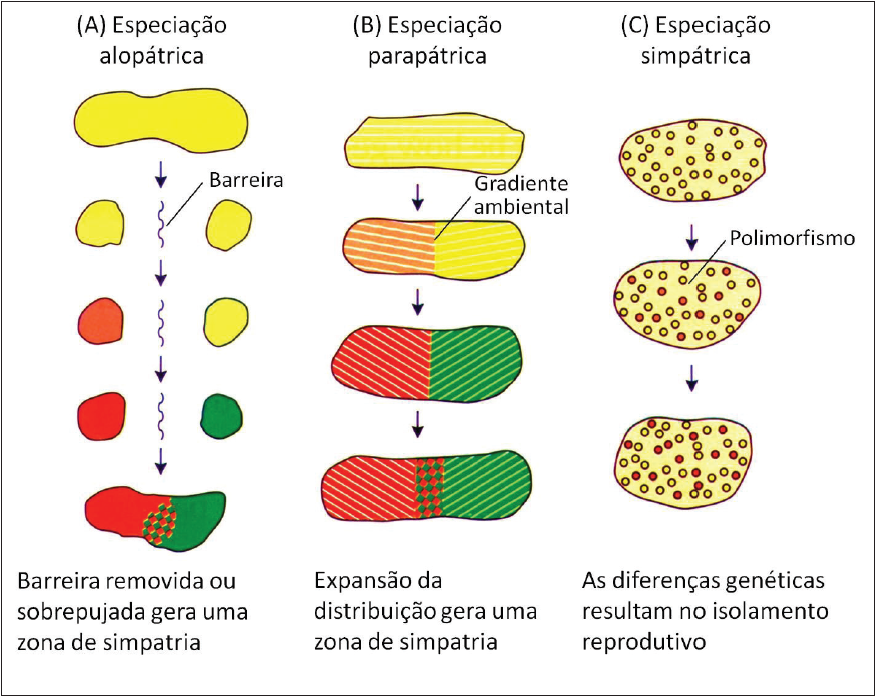
Figura 3.
Estágios dos três modelos de especiação de acordo com sua configuração geográfica.
A) Especiação alopátrica.
B) Especiação parapátrica.
C) Especiação simpátrica.
(Adaptado de Futuyma, 2005).
De acordo com o que vimos, tanto a deriva genética quanto a seleção natural podem gerar isolamento reprodutivo como um subproduto da diferenciação entre populações geograficamente afastadas. Mas, a seleção natural não poderia favorecer diretamente o isolamento reprodutivo? Aqui, cabe fazer um parêntese: as duas etapas da especiação vistas até agora – isolamento geográfico e o surgimento do isolamento reprodutivo como um subproduto – são visões estabelecidas há bastante tempo na teoria evolutiva. Porém, há alternativas para estes processos, as quais consistem em conceitos que começaram a ser evidenciados e aceitos mais recentemente pelos biólogos. São eles: especiação sem isolamento geográfico e seleção natural para isolamento reprodutivo.
Respondendo à nossa pergunta feita anteriormente (se a seleção natural poderia favorecer diretamente o isolamento reprodutivo) podemos introduzir o segundo dos conceitos, isto é, seleção para isolamento. Imagine duas das subespécies de Heliconius erato. Os eventuais híbridos formados em zonas de contato entre as populações podem apresentar um padrão de coloração misto que os predadores locais não aprenderam a evitar. Se os híbridos forem menos viáveis devido a esta perda da coloração defensiva, como deverá atuar a seleção natural? A menor chance de sobrevivência dos híbridos resulta na menor propagação dos genes que promovem este tipo de cruzamento. A seleção natural, portanto, favorecerá os indivíduos portadores de genes que promovem o outro tipo de cruzamento, isto é, entre aqueles que possuem o mesmo padrão mimético. Se esta seleção resultar em critérios de preferência e/ou sinais de reconhecimento específico cada vez mais eficazes, estas subespécies podem se isolar permanentemente, completando o processo de especiação. Esta seleção para o isolamento reprodutivo, portanto, reforça a divergência
iniciada pelo isolamento geográfico, de modo que este processo é justamente denominado reforço.
A A esta altura podemos questionar a importância do isolamento geográfico como primeira etapa da especiação, já que as subespécies de Heliconius não estavam mais isoladas por nenhuma barreira física. Aqui, portanto, entra a segunda área mencionada anteriormente como menos evidenciada: a especiação sem isolamento geográfico. Quando a população ancestral ocupava uma área adjacente àquela onde surgiu a nova espécie (como no caso de Heliconius), diz-se que a especiação
foi parapátrica (para = ao lado) (Figura 3B). Gradientes de variação no ambiente, como o tipo de solo, por exemplo, também podem levar a uma divergência adaptativa entre populações adjacentes e desencadear a especiaçãoparapátricapartir do mesmo cenário inicial imaginado, suponha que os híbridos sejam viáveis e prefiram
acasalar com seus semelhantes de coloração mista. O que aconteceria? Novamente o reforço estará ocorrendo, mas neste caso poderá surgir uma terceira espécie. Na verdade, este cenário não é fictício, já existindo um caso documentado de especiação por hibridização em Heliconius, entre H. cydno e H. melpomene, originando H. heuripa (MAVÁREZ et al., 2006). Se analisarmos melhor este exemplo, notamos que, na última etapa da divergência, a seleção sexual, através da preferência diferencial por parceiros, foi o fator evolutivo mais importante. Assim, não só a seleção natural, mas também a seleção sexual pode participar da especiação.
Em outro cenário, a nova espécie pode surgir no mesmo local de sua espécie ancestral em um processo denominado especiação simpátrica (sim = igual) (Figura 3C). Imagine que em uma população de moscas que utilizam o fruto “A” para oviposição apareça um polimorfismo genético que dá aos seus portadores a capacidade e a preferência para explorar um novo fruto “B”. Se as moscas macho procuram por suas parceiras nos locais de oviposição, é provável que haja um maior número de cruzamento entre os portadores da nova variação genética do que com os demais, assim como entre os indivíduos que exploramo fruto “A”. O fator importante neste, assim como em todos os exemplos de especiação, é a redução do fluxo gênico; com ou sem isolamento geográfico.
Como as espécies se mantém?
Uma vez que duas espécies já tenham se diferenciado, os fatores que as mantêm reprodutivamente isoladas são necessariamente os mesmos que as levaram às suas origens? Uma questão mais específica neste contexto é: o que acontece se as populações que divergiram recentemente têm a oportunidade de intercruzar? O contato secundário entre écies pode ter algumas consequências evolutivas importantes e uma delas já foi brevemente discutida neste artigo: a hibridização. Assim, se os híbridos tiverem menor aptidão do que as formas parentais, o reforço entra em jogo, aumentando uma divergência já iniciada. O reforço, portanto, pode atuar tanto durante quanto após um evento de especiação.
O reforço pode levar ao surgimento de “barreiras” que impedem ou diminuem significativamente as chances de hibridização. É importante mencionar, contudo, que estas barreiras também podem surgir por diversos outros caminhos além do reforço, dependendo da história evolutiva das espécies. Elas são chamadas de mecanismos de isolamento pré-zigóticos e podem atuar nos seguintes níveis:
- ecológico, quando os nichos diferem;
- sazonal, quando os períodos reprodutivos diferem;
- etológico, quando os sinais de reconhecimento diferem;
- fisiológico, quando hormônios importantes são específicos;
- mecânico, quando as genitálias não se encaixam por diferenças morfológicas;
- gamético, quando os gametas de um sexo são incompatíveis e resultam na morte dos do sexo oposto.
No entanto, as populações também podem permanecer geneticamente isoladas na ausência de reforço se a prole híbrida for inviável. Este tipo de barreira é denominado isolamento pós-zigótico e pode atuar no nível da mortalidade zigótica, embrionária ou do adulto, se ele for incapaz de sobreviver e ou de se reproduzir.
Os fatores envolvidos na especiação, conforme vistos nesta síntese, muitas vezes atuam simultaneamente e em mais de uma etapa do processo. O uso de conceitos acilita acompreensão e a interpretação de padrões observados na natureza, mas temos que nos manter cientes de que estes modelos teóricos constituem simplificações de algo uito ais complexo e intrincado.
Referências
COSTA-SCHMIDT, L. E.; ARAÚJO, A. M. Genital Variation and Taxonomic Discrimination in the Semi-Aquatic Spider Genus Paratrechalea (Araneae; Trechaleidae). The Journal of Arachnology v. 38, p. 242-249, 2010.
DODD, D. M. B. Reproductive isolation as a consequence of adaptive divergence in Drosophila pseudoobscura. Evolution v.43, p. 1308–1311, 1989.
FREEMAN, S.; HERRON, J. C. Análise Evolutiva. 4a ed. Porto Alegre: Artmed, 2009, 848 p. FUTUYMA, D. J. Evolution. 3a ed. Massachusetts: Sinauer Associates, Sunderland, 2005, 763 p.
HOWARD, D.; BERLOCHER, S. Endless forms: species and speciation. New York: Oxford University Press, 1998, 470 p. JIGGINS, C. D.. Ecological speciation in mimetic butterflies. BioScience v.58, n.6, p. 541- 548, 2008.
RIDLEY, M. Evolução. 3a ed. Porto Alegre: Artmed, 2004, 752 p.
MAVÁREZ, J.; SALAZAR, C.A.; BIRMINGHAM, E.; SALCEDO, C.; JIGGINS, C. D.; LINARES, M. Speciation by hybridization in Heliconius butterflies. Nature v.441, p. 68-871, 2006.
MAYR, E. Animal Species and Evolution. Cambridge: Harvard University Press, 1963 PPARD, P. M.; TURNER, J. R. G.; BROWN,
K. S.; BENSON, W. W.; SINGER, M. C. Genetics and the evolution of müllerian mimicry in Heliconius butterflies. Philosophical Transactions of the Royal Society of London (B) v.308, p. 433-613, 1985.
SILVA, L. M.; ARAÚJO, A. M. The genetic structure of Heliconius erato populations (Lepidoptera; Nymphalidae). Revista Brasileira de Genética v.17, n.1, p. 19-24, 1994.
WEINER J. O Bico do Tentilhão – Uma História da Evolução no Nosso Tempo. Rio de Janeiro: Editora Rocco, 1995.
André Luis Klein
Doutorando no Programa de Pós-Graduação em Genética e Biologia Molecular. Universidade Federal do Rio Grande do Sul, Departamento de Genética.
Claiton Martins-Ferreira
Pesquisador Pós-doc no Programa de Pós-Graduação em Genética e Biologia Molecular. Universidade Federal do Rio Grande do Sul, Departamento de Genética.
Endereço para correspondência: ndrklein@gmail.com
Como citar:
KLEIN, A. L. e MARTINS-FERREIRA, C.. Espécies: o que são e como surgem? Genética na Escola, on-line, São Paulo, v.7, n.2, 2012. Disponível: http://www.geneticanaescola.com.br/#!edicoes-anteriores/cudb . Acesso em [Dia] [Mês] [Ano]. ISSN 1980-3540.

Mauricio, agora que vi que nosso artigo está aqui. Agradeço mas tenho uma solicitação a fazer. Por favor, coloca a referência completa para quem usar esse artigo, pois do jeito que está não tem como citar. Obrigado.
CurtirCurtir